Støtter de vitenskapelige bevisene evolusjonære modeller av menneskelig opprinnelse?
Casey Luskin; 12. juni 2023 Oversatt herfra {Kursiv og understreking ved oversetter.}
Forfatterens notat: I et tidligere innlegg introduserte jeg artikkelen min, "Sammenligning av moderne evangeliske modeller om menneskelig opprinnelse," åpen tilgang og nylig publisert i det fagfellevurderte tidsskriftet Religions -lenke. Artikkelen gjennomgår åtte modeller for å forene vitenskap og kristen teologi angående menneskelig opprinnelse. Fire av disse modellene er evolusjonære: (1) den klassiske teistiske evolusjons-/evolusjons kreasjonisme-modellen, (2) Homo divinus-modellen, (3) Den genealogiske Adam og Eva-modellen og (4) Homo heidelbergensis-modellen. Og fire av disse modellene anses som ikke-evolusjonære: (5) Unik opprinnelse Design-modellen, (6) Klassisk gammel jord kreasjonist-modellen, (7) Klassisk ung jord Kreasjonist-modellen og (8) Gammel jord/Nåværende mennesker hybrid modellen.
Artikkelen går langt utover intelligent design og kommer inn på et teologisk territorium som er noe fremmed for meg og forskjellig fra mitt typiske fokus på vitenskap. Men det er også mye vitenskap diskutert i artikkelen. I den forbindelse vil jeg nedenfor gi et utdrag fra en del av artikkelen der jeg diskuterer hvordan den teistiske evolusjon/evolusjonære skapelsen (TE/EC)-modellen holder seg i lys av de vitenskapelige bevisene. Selv om materialet nedenfor er innrammet som et svar på TE/EC-modellen, er mye av det vitenskapelige beviset som er reist også anvendelig for å vurdere hvilken av de fire evolusjonsmodellene som jeg vurderer i artikkelen.
En åpenbar fordel med [Teistisk Evolusjon / Evolutionær kreasjinisme]-modellen er at den er fullt kompatibel med mainstream evolusjonsvitenskap. Men ved å benekte eksistensen av en historisk Adam og Eva, avviser den eksplisitt store tradisjonelle teologiske oppfatninger om menneskelig opprinnelse, noe som ikke vil være akseptabelt for mange som søker å bevare denne troen (Grudem 2017a, 2017b). Før de aksepterer TE/EC-modellen, kan de som verdsetter disse store tradisjonelle teologiske overbevisningene først vurdere i hvilken grad vitenskapelige bevis mot eksistensen av Adam og Eva og til fordel for menneskelig evolusjon, er så klare og opplagte som det hevdes.
Bilde 1. Forskjell mellom teori og empiri
 Fossile bevis
Fossile bevis
For det første viser fossilregistreret et tydelig brudd mellom de apelignende australopithecines, som visstnok er direkte forfedre til vår slekt Homo, og de første menneskelignende medlemmene av slekten Homo (Luskin 2022). Slike bevis har ført til observasjoner fra hovedstrøms-evolusjonister som innrømmer at det er et "stort gap uten bro" mellom menneskelignende medlemmer av Homo og australopithecinene (Mayr 2004, s. 198), som krevde en "genetisk revolusjon" siden "ingen australopithecin-arter er åpenbart overgangsbestemt" (Hawks et al. 2000, s. 4), og innebærer en "big bang"-modell av menneskelig opprinnelse (University of Michigan News Service 2000). Mens evolusjonære paleoantropologer generelt mener at "overgangen fra Australopithecus til Homo utvilsomt var en av de mest kritiske i sitt omfang og konsekvenser," innrømmer de at "mange detaljer om denne overgangen er uklare på grunn av mangelen på fossile og arkeologiske registreringer" ( Lieberman et al. 2009, s. 1). Denne mangelen på fossile bevis for utviklingen av den menneskelignende kroppsplanen i fossilregisteret svekker nødvendigheten av å ta i bruk standard evolusjonære forklaringer av menneskelig opprinnelse.
 Genetisk bevis
Genetisk bevis
Genomiske sammenligninger mellom mennesker og sjimpanser blir mer sofistikerte, med nyere forslag som foreslår lavere estimater av menneske-sjimpans genetiske likhetsestimater til mellom 84 prosent og 96 prosent (Buggs 2018c; Seaman og Buggs 2020). Det er også uklart hvordan noen prosent likhet mellom menneskelig og sjimpanse-DNA gir et argument som krever felles aner (Luskin 2022). (Se referanser i originalartikkelen.)
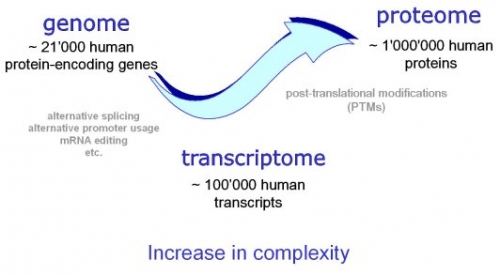
Bilde 2. Menneskelig genom -transkriptom-proteom
Junk-DNA-genetiske argumenter for felles menneske-ape-forfedre har også blitt kritisert i de siste årene på grunn av oppdagelsen av massefunksjonalitet for ikke-kodende eller "søppel" DNA i det menneskelige genomet. En stor Nature-artikkel fra 2012 av ENCODE-konsortiet rapporterte "biokjemiske funksjoner for 80 %" av det menneskelige genomet (ENCODE Project Consortium 2012, s. 57). Ledende ENCODE-forskere spådde at med videre forskning vil "80 prosent gå til 100" siden "nesten hvert nukleotid er assosiert med en funksjon." (Yong 2012). I kjølvannet av denne forskningen publiserte tidsskriftet Science en artikkel med tittelen "ENCODE Project Writes Eulogy for Junk DNA" som uttalte at disse funnene "innebærer dødsstøtet for ideen om at vårt DNA stort sett er strødd med ubrukelige baser" (Pennisi 2012, s. 1159). Bevis på funksjoner for ikke-kodende DNA har fortsatt å øke i høyt tempo. En artikkel i Nature fra 2021 rapporterte at over 130 000 spesifikke "genomiske elementer, tidligere kalt søppel-DNA" har sett spesifikke funksjoner identifisert (Gates et al. 2021, s. 215), etterfulgt av en artikkel i Genome Biology and Evolution som konkluderte, "Tiden for 'søppel-DNA' er over" (Stitz et al. 2021, s. 11). Det er fortsatt mye vi ikke forstår om genomet, og det er mange spesifikke genetiske elementer som ingen funksjon ennå er oppdaget for. Ikke desto mindre antyder dette beviset en sterk trendlinje i forsknings-litteraturen, bort fra ikke-funksjonalitet for "søppel" DNA.
 Et ofte nevnt eksempel på søppel-DNA er pseudogener, som TE/EC-forkjempere ofte siterer som "de muterte restene av gener som en gang var funksjonelle" (Venema og Falk 2010) som viser vår felles aner med aper. Likevel fant en artikkel fra 2012 at pseudogen funksjon er "utbredt", og siden "studiet av funksjonelle pseudogener bare er i begynnelsen" spådde det "flere og flere funksjonelle pseudogener vil bli oppdaget etter hvert som nye biologiske teknologier utvikles" (Wen et al. 2012, s. 31). Faktisk er litteraturen nå full av artikler som rapporterer funksjoner for pseudogener (Hirotsune et al. 2003; Pain et al. 2005; Zhang et al. 2006; Tam et al. 2008; Piehler et al. 2008; Poliseno et al. 2010; Muro et al. 2011; Rapicavoli et al. 2013; Ji et al. 2015; Hayashi et al. 2015; Prieto-Godino et al. 2016; Suzuki et al. 2018; Fiddes et al. 2018; Habib et al. 2019) , inkludert å produsere funksjonelle RNA-transkripsjoner eller utføre funksjoner uten å produsere noe RNA (Poliseno 2012). En artikkel fra 2012 bemerker at selv om "pseudogener lenge har blitt avvist som søppel-DNA", har nyere fremskritt fastslått at "DNA til et pseudogen, RNA transkribert fra et pseudogen, eller proteinet oversatt fra et pseudogen kan ha flere, forskjellige funksjoner," som konkluderer med at "pseudogener har dukket opp som en tidligere ikke verdsatt klasse av sofistikerte modulatorer av genuttrykk" (Poliseno 2012, s. 1, 10). Faktisk er mange pseudogener kjent for å produsere proteiner, med en studie i Nature som rapporterte "mer enn 200 peptider, som er kodet av 140 pseudogener" (Kim et al. 2014, s. 579).
Et ofte nevnt eksempel på søppel-DNA er pseudogener, som TE/EC-forkjempere ofte siterer som "de muterte restene av gener som en gang var funksjonelle" (Venema og Falk 2010) som viser vår felles aner med aper. Likevel fant en artikkel fra 2012 at pseudogen funksjon er "utbredt", og siden "studiet av funksjonelle pseudogener bare er i begynnelsen" spådde det "flere og flere funksjonelle pseudogener vil bli oppdaget etter hvert som nye biologiske teknologier utvikles" (Wen et al. 2012, s. 31). Faktisk er litteraturen nå full av artikler som rapporterer funksjoner for pseudogener (Hirotsune et al. 2003; Pain et al. 2005; Zhang et al. 2006; Tam et al. 2008; Piehler et al. 2008; Poliseno et al. 2010; Muro et al. 2011; Rapicavoli et al. 2013; Ji et al. 2015; Hayashi et al. 2015; Prieto-Godino et al. 2016; Suzuki et al. 2018; Fiddes et al. 2018; Habib et al. 2019) , inkludert å produsere funksjonelle RNA-transkripsjoner eller utføre funksjoner uten å produsere noe RNA (Poliseno 2012). En artikkel fra 2012 bemerker at selv om "pseudogener lenge har blitt avvist som søppel-DNA", har nyere fremskritt fastslått at "DNA til et pseudogen, RNA transkribert fra et pseudogen, eller proteinet oversatt fra et pseudogen kan ha flere, forskjellige funksjoner," som konkluderer med at "pseudogener har dukket opp som en tidligere ikke verdsatt klasse av sofistikerte modulatorer av genuttrykk" (Poliseno 2012, s. 1, 10). Faktisk er mange pseudogener kjent for å produsere proteiner, med en studie i Nature som rapporterte "mer enn 200 peptider, som er kodet av 140 pseudogener" (Kim et al. 2014, s. 579).
Bilde 3. Forskjell på god og dårlig (pseudo)vitenskap
Det er gode grunner til å forstå hvorfor DNA merket som "pseudogen" nå viser seg å ha en funksjon. Pseudogener blir ofte identifisert ved sammenligning med noen andre lignende gensekvenser som koder for et funksjonelt protein som pseudogenet antas å ha blitt avledet fra og deretter degenerert. Imidlertid krever regulatoriske funksjoner til pseudogener ofte at de har en viss sekvenslikhet (homologi) med deres proteinkodende motstykker, slik at RNA-transkriptene deres kan samhandle med transkripsjoner fra det proteinkodende genet, og derved regulere proteinproduksjonen (Salmena et al. 2011). Derfor er grunnen til at genomene våre inneholder sekvenser som ligner proteinkodende gener, men som ikke produserer proteiner, ikke fordi de er forkastet evolusjonære "pseudogener", men fordi de er utformet på den måten, som viktige genomiske regulerings- og kontrollelementer.
Mange artikler har kritisert antakelsen om at pseudogener er funksjonsløst søppel-DNA og foreslår nå å forlate begrepet (Pink et al. 2011; Poliseno 2012; Wen et al. 2012; Kovalenko og Patrushev 2018; Troskie et al. 2021). En artikkel fra 2020 i Nature Reviews Genetics observerer: "Der pseudogener har blitt studert direkte, er de ofte funnet å ha kvantifiserbare biologiske roller," og advarer om at "den dominerende begrensningen i å fremme etterforskningen av pseudogener nå ligger i fellene til den rådende tankegangen at pseudogene regioner i seg selv er ikke-funksjonelle." Den advarer mot at pseudogen funksjon er "for tidlig avvist" på grunn av "dogmer" (Cheetham et al. 2020).
Det er fremtredende eksempler på for tidlig avvisning av pseudogen funksjon, for så å bli bevist feil senere. Under Kitzmiller mot Dover-rettssaken i 2005, vitnet biolog Kenneth Miller om at vårt beta-globin-pseudogen er "ødelagt" fordi det har "molekylære feil som gjør genet ikke-funksjonelt", noe som indikerer at mennesker deler en felles stamfar med aper (Miller 2005, s. 79). To år senere hevdet den ledende evolusjonsforkjemperen Eugenie Scott at dette pseudogenet "ikke kommer til å gjøre noe vanskelig. Det kommer bare til å sitte der" og "ikke gjøre noe" (Scott 2007). En studie fra 2013 rapporterte imidlertid at presist dette pseudogenet er funksjonelt (Moleirinho et al. 2013), og en studie fra 2021 fant at det er "essensielt" og er "uunnværlig" for dannelse av røde blodlegemer (Ma et al. 2021, s. 478, 490).
 Endogene retrovirus (ERV-er) er en annen klasse av "søppel"-DNA som ofte siteres av TE/EC-er til fordel for menneskers/apers felles aner. Strukturen til argumentene her er lik de som brukes for pseudogener: ERV-er viser også utbredt bevis på funksjon, spesielt genregulerende roller (Conley et al. 2008; Jacques et al. 2013; Chuong 2018; Robson og Mundlos 2019; Jönsson et al. . 2021; Sakashita et al. 2023), og deres likhet med viralt DNA kan eksistere av funksjonelle årsaker, ettersom de er involvert i immunfunksjoner for å avvise virusinfeksjoner (Badarinarayan og Sauter 2021).
Endogene retrovirus (ERV-er) er en annen klasse av "søppel"-DNA som ofte siteres av TE/EC-er til fordel for menneskers/apers felles aner. Strukturen til argumentene her er lik de som brukes for pseudogener: ERV-er viser også utbredt bevis på funksjon, spesielt genregulerende roller (Conley et al. 2008; Jacques et al. 2013; Chuong 2018; Robson og Mundlos 2019; Jönsson et al. . 2021; Sakashita et al. 2023), og deres likhet med viralt DNA kan eksistere av funksjonelle årsaker, ettersom de er involvert i immunfunksjoner for å avvise virusinfeksjoner (Badarinarayan og Sauter 2021).
Bilde 4. ERVer kan omforme RNA
Igjen er det sant at det fortsatt er mye vi ikke vet om søppel-DNA, og det er mange spesifikke genetiske elementer (inkludert pseudogener og ERV-er) som spesifikke funksjoner ennå ikke er oppdaget for. Nyere trender innen forskning viser imidlertid at langt mer funksjonalitet blir oppdaget enn forventet, noe som fører til muligheten for massefunksjonalitet for søppel-DNA. Som en akademisk bok fra 2023 om RNA sier:
Mens historien fortsatt utfolder seg, konkluderer vi med at genomene til mennesker og andre komplekse organismer ikke er fulle av søppel, men snarere er svært kompakte informasjonspakker som i stor grad er viet til spesifikasjonen av regulatoriske RNA-er. Disse RNA-ene driver banene for differensiering og utvikling, underbygger hjernefunksjonen og formidler transgenerasjonsminne av erfaring, mye av det i strid med langvarige forestillinger om genetisk programmering og evolusjonsteoriens dogmer. -MATTICK OG AMARAL 2023, S. VII.
Hvis ikke-kodende/søppel-DNA faktisk er funksjonelt, kan genetiske likheter være et resultat av felles design på grunn av behovet for å oppfylle lignende funksjonskrav. Til og med Francis Collins har erkjent at delt genetisk likhet "alene, selvfølgelig, ikke beviser en felles stamfar" fordi "slike likheter ganske enkelt kunne demonstrere at Gud brukte vellykkede designprinsipper om og om igjen" (Collins 2006, s. 134).
Populasjonsgenetikk
 For det tredje synes argumenter mot Adam og Eva basert på menneskelig genetisk mangfold og populasjonsgenetikk å ha blitt undergravd av påfølgende modelleringsanalyser. En av de første vitenskapelige svarene på dette populasjonsgenetikk-argumentet mot Adam og Eva ble publisert i 2012, da biolog Ann Gauger, seniorstipendiat ved Discovery Institute (en pro-intelligent-design tenketank, hvor denne forfatteren også jobber), rapporterte at genetisk mangfold i HLA-gener - noen av de mest forskjellige genene i det menneskelige genomet - fortsatt kunne forklares hvis vi stammer fra et første par (Gauger et al. 2012, s. 120).
For det tredje synes argumenter mot Adam og Eva basert på menneskelig genetisk mangfold og populasjonsgenetikk å ha blitt undergravd av påfølgende modelleringsanalyser. En av de første vitenskapelige svarene på dette populasjonsgenetikk-argumentet mot Adam og Eva ble publisert i 2012, da biolog Ann Gauger, seniorstipendiat ved Discovery Institute (en pro-intelligent-design tenketank, hvor denne forfatteren også jobber), rapporterte at genetisk mangfold i HLA-gener - noen av de mest forskjellige genene i det menneskelige genomet - fortsatt kunne forklares hvis vi stammer fra et første par (Gauger et al. 2012, s. 120).
Bilde 5. Definisjon av populasjon (generelt)
Etter publiseringen av Adam and the Genome i 2017, ble biolog Dennis Venema engasjert på BioLogos diskusjonsforum av Richard Buggs, en genetiker ved Queen Mary University London. Under diskusjonen erkjente Venema at forskjellige artikler han hadde sitert for å ha tilbakevist eksistensen av Adam og Eva, faktisk ikke hadde tatt opp spørsmålet om menneskeheten stammet fra et første par (Buggs 2017). Fra denne diskusjonen oppsto en konsensus blant forskjellige kristne biologer om at hvis et første par mennesker levde langt nok i fortiden, så kunne moderne menneskelig genetisk mangfold faktisk forklares av naturlige biologiske prosesser. Til slutt uttalte Buggs til Venema: "Du ville gjort leserne dine en tjeneste hvis du skrev en blogg for å fortelle dem nå, så langt du kan, at dagens genomiske mangfold hos mennesker ikke utelukker en flaskehals i den menneskelige avstamningen mellom ca. 700 tusen og 7mill. år siden. Jeg tror du skylder dem dette" (Buggs 2018b). (Flaskehalsen han refererer til, er at den menneskelige befolkningen blir redusert til to individer - i praksis det samme som at menneskeheten stammer fra Adam og Eva.) Venema erkjente da offentlig sannheten av Buggs kritikk, og svarte: "Jeg har allerede vært enig i dette .. Du må gjerne offentliggjøre det" (Venema 2018). Buggs oppsummerte senere denne samtalen på bloggen sin med Nature Ecology and Evolution, og rapporterte at spørsmålet ikke lenger var om en historisk Adam og Eva kunne ha eksistert, men på hvilket tidspunkt de levde (Buggs 2018a).
 Gauger, sammen med Ola Hössjer, professor i matematikk ved Universitetet i Stockholm, hadde allerede startet et prosjekt for å ta for seg hvor dypt inn i fortiden et første par måtte leve for å ta hensyn til moderne menneskelig genetisk mangfold. De publiserte en serie artikler som utviklet og tester en populasjonsgenetikkmodell som gjør det mulig for et første par mennesker å bli gitt "designede varianter" av gener som representerer "urmangfold" innebygd i de opprinnelige genomene til Adam og Eva. Under deres modell styrer så naturlige biologiske prosesser menneskehetens påfølgende genetiske historie. De viste at moderne menneskelig genetisk mangfold kan forklares av et enkelt par forfedre - for eksempel det man kan kalle Adam og Eva - forutsatt at de levde for minst 500 000 år siden (Hössjer et al. 2016a, 2016b; Hössjer og Gauger 2019). S. Joshua Swamidass, en kristen forsker og professor i beregningsbiologi ved Washington University i St. Louis, utførte en analyse som ga et lignende resultat, og fant ut at Adam og Eva kunne ha levd for 495 000 år siden som våre eneste genetiske stamfedre (Swamidass 2017).
Gauger, sammen med Ola Hössjer, professor i matematikk ved Universitetet i Stockholm, hadde allerede startet et prosjekt for å ta for seg hvor dypt inn i fortiden et første par måtte leve for å ta hensyn til moderne menneskelig genetisk mangfold. De publiserte en serie artikler som utviklet og tester en populasjonsgenetikkmodell som gjør det mulig for et første par mennesker å bli gitt "designede varianter" av gener som representerer "urmangfold" innebygd i de opprinnelige genomene til Adam og Eva. Under deres modell styrer så naturlige biologiske prosesser menneskehetens påfølgende genetiske historie. De viste at moderne menneskelig genetisk mangfold kan forklares av et enkelt par forfedre - for eksempel det man kan kalle Adam og Eva - forutsatt at de levde for minst 500 000 år siden (Hössjer et al. 2016a, 2016b; Hössjer og Gauger 2019). S. Joshua Swamidass, en kristen forsker og professor i beregningsbiologi ved Washington University i St. Louis, utførte en analyse som ga et lignende resultat, og fant ut at Adam og Eva kunne ha levd for 495 000 år siden som våre eneste genetiske stamfedre (Swamidass 2017).
Bilde 6. Ventetidsproblemet generelt (G. Bechly)
Det er et annet populasjonsgenetikkargument som er relevant for menneskelig opprinnelse - men dette er stilt som en matematisk utfordring for ikke-styrte evolusjons-modeller. MRCA for mennesker og sjimpanser sies å ha levd for omtrent 4 til 6 millioner år siden (Wood og Harrison 2011). Selv om estimatene varierer, foreslår en rimelig beregning at de genetiske forskjellene mellom mennesker og sjimpanser utgjør rundt 35 millioner baseparendringer, 5 millioner indels [sekvenser av flere nukleotidbaser] i hver art, og 689 ekstra gener hos mennesker" (Cohen) 2007). Likevel fant en populasjonsgenetikkstudie i tidsskriftet Genetics, at hvis bare to spesifikke mutasjoner var nødvendig for å gi en evolusjonær fordel i linjen som førte til mennesker, så ville "denne typen endring ta >100 millioner år", som ble bestemt til å være "svært usannsynlig å finne sted på en rimelig tidsskala" (Durrett og Schmidt 2008). Dette "ventetids"-problemet (Hössjer et al. 2021) antyder at det kan være alt for lite tid tilgjengelig fra fossilregistrene for standard ikke-styrte evolusjonsmekanismer til å generere observerte genetiske, morfologiske og atferdsmessige forskjeller mellom mennesker og sjimpanser. Noen har foreslått intelligent design som en mulig forklaring på den raske opptredenen av biologisk informasjon som er nødvendig, for å overvinne ventetidsproblemet og generere disse komplekse egenskapene (Thorvaldsen og Hössjer 2020).
Igjen, hele artikkelen er åpen og tilgjengelig her.
Oversettelse via google oversetter og bilder ved Asbjørn E. Lund